ブドウ糖から脂肪酸をつくる時、アセチルCoAからスタートして、まず炭素数16のパルミチン酸をつくります。もともとアセチルCoAからつくられた、アセチルACPとマロニルACPを加えてできた炭素数4のブチリルACPに、マロニルACPから炭素を2個ずつ足して炭素数16のパルミチン酸まで延ばします。
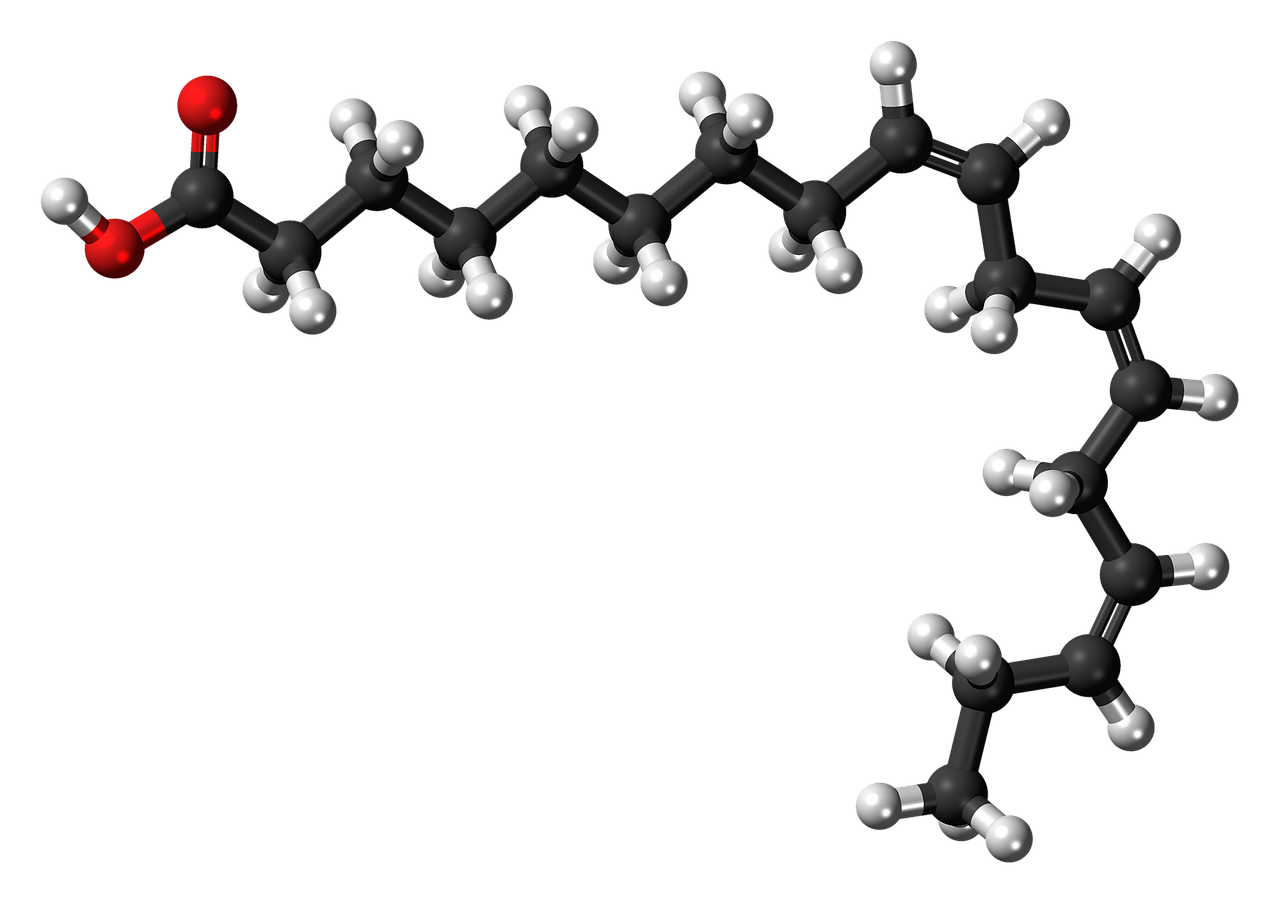
脂肪は脂肪酸とグリセリン(グリセロール)に分解されます。脂肪酸もグリセリンも体の中で再び脂肪となりますが、「糖分をとり過ぎると太る」といわれるように、ブドウ糖から脂肪がつくられる仕組みもあります。
以前、ブドウ糖を分解する解糖系から脂肪合成に必要なグリセリンを得るという記事を書いて、解糖系の中でできるジヒドロキシアセトンリン酸をもとに、リン酸がついた「活性化」したグリセリンであるグリセロール3-リン酸ができることがわかりました。
この記事では、脂肪を構成するもう一つのパーツである脂肪酸について、ブドウ糖から生合成される仕組みを説明します。
パルミチン酸をまずつくる
脂肪酸を生合成する場合、炭素数16(C:16)の飽和脂肪酸パルミチン酸をまずつくります。他の脂肪酸は、出来上がったパルミチン酸を利用してつくられます。
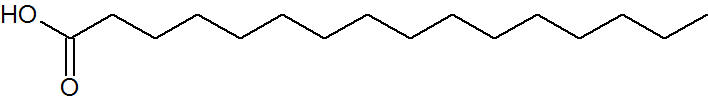
パルミチン酸
脂肪酸生合成
脂肪酸から炭素2個ずつ切り出して短くしていくβ酸化はミトコンドリア内で行われましたが、逆に炭素を2個ずつつなげて脂肪酸をつくる生合成は、ミトコンドリアの外、細胞質ゾルで行われます。
しかし、出発点は、オキサロ酢酸とアセチルCoAです。TCA回路のスタートと同じです。クエン酸ができます。このクエン酸をミトコンドリアの外に出してスタートします。
アセチルCoAとオキサロ酢酸からクエン酸
まず、ミトコンドリアの中で解糖系ピルビン酸由来のアセチルCoAとオキサロ酢酸からクエン酸が作られます。クエン酸はミトコンドリアの中から外の細胞質ゾルへ出ていくことができるので、細胞質ゾルで分解されて再びアセチルCoAとなります。
アセチルCoAはミトコンドリアの外に出られない
イラストレイテッド ハーパー・生化学原書29版にはこのように書かれています。
アセチル-CoAは,グルコース由来のピルビン酸がミトコンドリアで酸化されることで生成する.しかしならが,アセチル-CoAは脂肪酸合成の主要な場である(ミトコンドリア外の)サイトゾルには容易に拡散できない.
クエン酸回路においてアセチル-CoAとオキサロ酢酸の縮合で生じたミトコンドリア内のクエン酸は,トリカルボン酸輸送体によりミトコンドリア外へ運び出され,そこでCoAとATPの存在下にATP-クエン酸リアーゼのはたらきで,アセチル-CoAとオキサロ酢酸に分解される.
さて、アセチルCoAは、クエン酸シンターゼという酵素の働きによってオキサロ酢酸と結合して、CoAが外れてクエン酸になります。
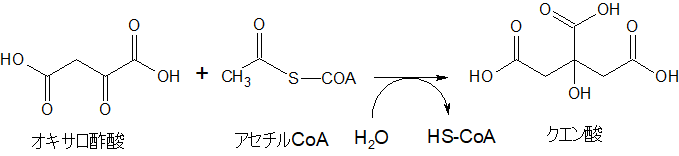
クエン酸は、ミトコンドリアの膜を通過することができます。つまり、クエン酸はミトコンドリアから外の細胞質ゾルに出ることができます。
ちなみに、アセチルCoAはミトコンドリアから外に出られないのだそうです。この辺は面白いところです。
クエン酸から再びアセチルCoAとオキサロ酢酸へ戻る
クエン酸は、細胞質ゾルで、ATP-クエン酸リアーゼという酵素によって、再び、CoAをつけて、アセチルCoAとオキサロ酢酸に戻ります。
KEGGでは、ATPクエン酸リアーゼについて、ENZYME: 2.3.3.8に説明と反応式があります。
こうして、ミトコンドリアから細胞質ゾルに実質的にアセチルCoAが出されて、脂肪酸の生合成が行われるようになります。
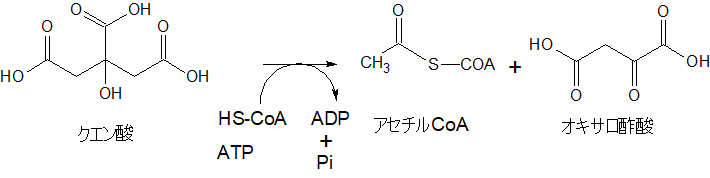
さて、ここから先は、まずパルミチン酸がどのように作られていくのかを見ていきます。
アセチルCoAからマロニルCoAへ
細胞質ゾルでは、ミトコンドリアから運ばれてきたアセチルCoAをまず、マロニルCoAという物質に変えます。
ここからは、KEGGのFatty acid biosynthesis(脂肪酸生合成)を読んで行きます。
この反応は、ENZYME: 6.4.1.2に詳しい説明があります。以下、同じように読み進めていきます。
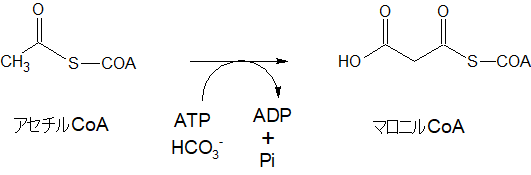
アセチルCoAは炭素数が2でしたが、マロニルCoAは、炭素数が3になっています。
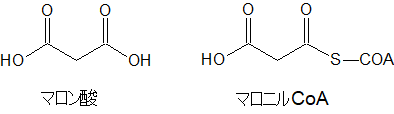
マロン酸
マロン酸に補酵素A(HS-CoA)がついたのがマロニルCoAです。マロン酸はカルボキシ基(COOH)が2個ついています。
脂肪酸はカルボキシ基が1個しかついていないので、そこが違いです。
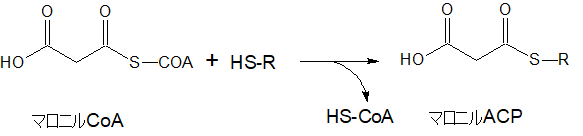
マロニルCoAからマロニルACPへ
さらに、マロニルCoAにアシルキャリアプロティン(ACP)というタンパク質に結合させると同時にCoAをはずして、マロニルACP(ACPはAcyl Carrier Protein)となります。
このマロニルACPが脂肪酸を作っていく上で最も重要な物質になります。炭素数を2個ずつ増やすもとになる物質です。
アシルキャリアプロティンは、アシルキャリアタンパク質ともいわれ、ACPと略されます。分子量10,000の小型タンパク質です。補酵素A(CoA)の分子量は757程度なのでACPの方がはるかに大きいです。
上の図では、HS-Rと表されています。
CoAと同じく-SH基を持ち、CoAと同じように自身は変化を受けずにアシル基を運ぶ役割を持ちます。キャリア(carrier)ですからね。そのまんまです。
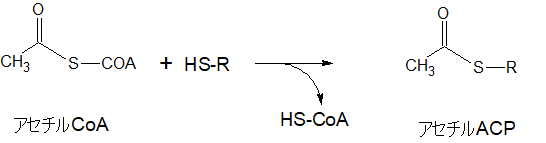
アセチルCoAからアセチルACPへ
一方で、別のアセチルCoAにACPが結合し、CoAが外れてアセチルACPになります。最初にマロニルACPが結合する物質です。
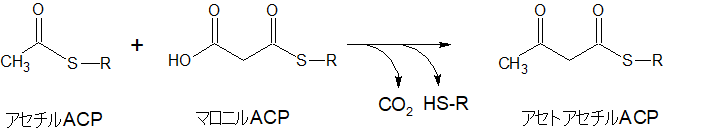
アセチルACPにマロニルACPが結合、アセトアセチルACPへ
アセチルACPにマロニルACPが結合し、ACP1個とCO2を放出し、アセトアセチルACPとなります。
炭素数は2+3-1=4
結果的に、もともと炭素数2だったアセチルACPがアセトアセチルACPへになることで炭素数が2個増えています。
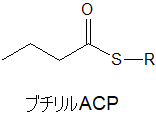
アセトアセチルACPからブチリルACPへ
アセトアセチルACPは、3つの反応を経て最終的に炭素数4のブチリルACPへ変化します。ブチリルACPは、見ての通り、ACPがついていなければ脂肪酸です。
ここからの反応は、余分な酸素(O)を外す反応です。炭素数は4のまま変化しません。
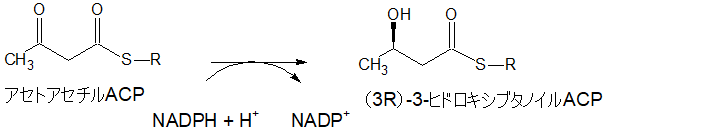
(3R)-3-ヒドロキシブタノイルACP
アセトアセチルACPは、補酵素NADPH+H+から水素を2個もらい、(3R)-3-ヒドロキシブタノイルACPになります。
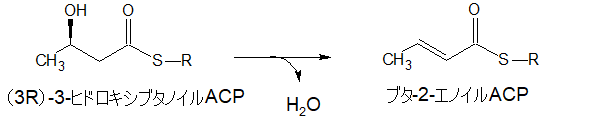
ブタ-2-エノイルACP
さらに脱水してブタ-2-エノイルACPになります。
ブチリルACP
ブタ-2-エノイルACPは補酵素補酵素NADPH+H+から水素を2個もらい、ブチリルACPとなります。
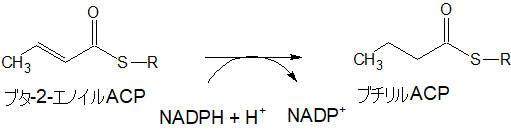
次の反応では、炭素数4のブチリルACPに炭素2個加えたヘキサノイルACPができます。
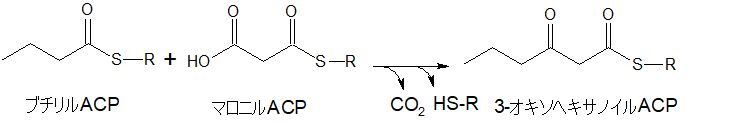
ブチリルACPから3-オキソヘキサノイルACPへ
ブチリルACPに、再び合成されてきたマロニルACPが結合します。
上と同じように、二酸化炭素を放出、炭素を1個減らしながら、実質炭素数を2個増やします。今までと同じ反応です。
(ブチリルACPの左端にCH3が書かれていないですが省略しているだけで同じブチリルACPです)
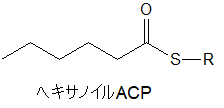
3-オキソヘキサノイルACPからヘキサノイルACPへ
3-オキソヘキサノイルACPは、同じように3つの酵素反応によって、炭素についている余分な酸素を取り外すとともに、炭素のすべての結合部分に水素を結合させて、ヘキサノイルACPとなります。
同様の反応を繰り返すことによってマロニルACPからの炭素を2個ずつ増やしていって、次第に長い炭素鎖のアシルACPができてきます。
パルミチン酸の完成
炭素数が16の長さになると、端のアシルキャリアプロティンが切れて、カルボキシル基となり、パルミチン酸が完成することになります。
β酸化は脂肪酸から炭素数が2個ずつ減っていきましたが、こちらは炭素数が2個ずつ増えていきます。
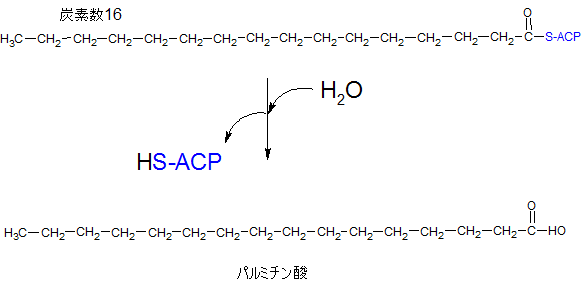
パルミチン酸完成
NOTE
ミトコンドリアの中で、アセチルCoAとオキサロ酢酸からクエン酸ができ、クエン酸がミトコンドリアの外の細胞質ゾルに出て来て、それを分解してアセチルCoAが得られます。
アセチルCoAの補酵素A(CoA)もそうですが、マロニルACPでは、運搬役に自身は変化を受けないACP(アシルキャリアプロティン)が登場します。ACPは、分子量10,000の小型タンパク質だと書かれていました。しかし、運搬されるのはわずか炭素数3の小さな物質です。
こういうアンバランスな仕組みが妙に面白いなと思ってしまいます。