脂肪が体の中でエネルギーとなるためには、脂肪から脂肪酸が切り離され、さらに分解されてアセチルCoAになる必要があります。
アセチルCoAをつくる行程は、脂肪酸からアシルCoAに変わった後、アシルCoAのβ位に二重結合ができ、二重結合に水が反応してOHが付き、OHがケト基(=O)に変わり、新たにHS-CoAが来てアセチルCoAが切り離されるという、4つの行程からなります。

脂肪酸が分解される場所は、ミトコンドリア。酸素呼吸の中枢の場所です。酸素呼吸では、まずブドウ糖を燃やすと説明されます。ブドウ糖はピルビン酸からアセチルCoAに分解されます。
脂肪酸もアセチルCoAまで分解されます。
β酸化、まず脂肪は脂肪酸とグリセリンになる
脂肪は、細胞の中でアシルCoAまで分解され、さらにアシルCoAはミトコンドリアの中でβ酸化という作用を受けてアセチルCoAになり、そしてTCA回路に入ります。
このあたりのことは私自身が詳しく知っておきたいので、詳しく調べながら書いていきます。
脂肪はまずリパーゼという酵素によって脂肪酸とグリセリンに分解されます。脂肪酸が次にアシルCoAになります。
アシル基
アシル基は脂肪酸からOH(ヒドロキシ基)を抜いたもののことをいいます。
アシルCoAのアシルとは、下の図を見てください。
パルミチン酸は炭素数16の飽和脂肪酸です。
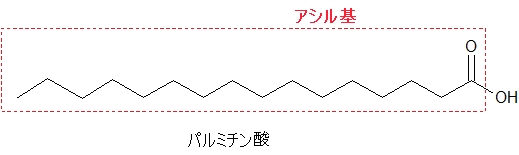
脂肪酸には必ずカルボキシ基(-COOH)がついています。脂肪酸からOH(ヒドロキシル基)を抜かした、赤い点線で囲まれた部分をアシル基といいます。
脂肪酸には炭素数、二重結合の数などいくつも種類がありますが、どんな脂肪酸でもOHを抜かしたものがアシル基です。
脂肪酸にCoAがついてアシルCoAになる
脂肪酸は、OH(ヒドロキシ基)部分で、補酵素A(HS-CoA)と結合し、アシルCoAとなります。
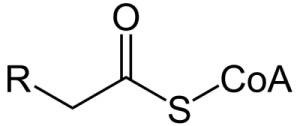
細胞の中(細胞質ゾル)で脂肪が分解されて切り離された脂肪酸は、補酵素A(HS-CoA)と結合してアシルCoAとなります。下図はアシルCoAの一般式です。
S-CoA以外が上で説明したアシル基です。Rは炭化水素鎖です。長さは脂肪酸によって違っていて何種類もありますから省略されてRになっています。S-CoAは補酵素Aです。アシル基とつながるCoAの末端の足がS(硫黄)です。
アシルCoAができるには、脂肪酸のOHと補酵素A(HS-CoA)のHが反応し、水(H2O)ができて離れ、それ以外が結合します。
アシルCoA
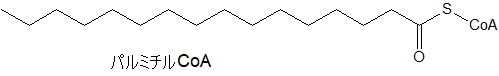
アシルCoAの一例として、炭素数16のパルミチルCoAを書いておきます。飽和脂肪酸のパルミチン酸に補酵素A(CoA)が結合したものです。
補酵素(HS-CoA)とは
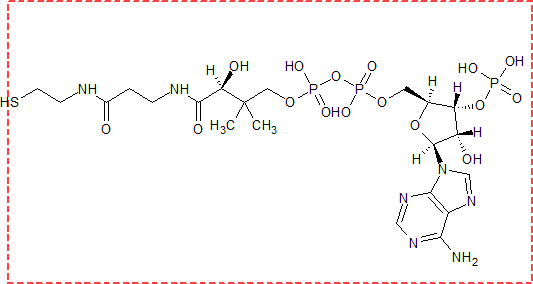
補酵素は、コエンザイム(coenzyme)とも呼ばれます。補酵素A(HS-CoA)の役割は、アシル基を運搬することです。補酵素A自体は反応を受けません。
補酵素A(以下CoAと書きます)とは、下図に書いた構造式のような物質です。赤い点線で囲まれている部分が、上の構造式にある、S-CoAの部分です。下図でH(水素)を赤い点線から外したのは、他のものと結合する時に外れてしまうからです。
しかし、CoAは補助分子という割には構造が複雑ですね。いままで見てきた脂肪酸の構造が簡単すぎるのでしょうか。
CoA
CoAの役割は、運搬することで自身は変化を受けません。そのため、このCoAの構造式を見る機会はあまりありません。
さて、脂肪酸にこの複雑なCoAがくっついたアシルCoAは、これからミトコンドリアに入ります。
アシルCoAはCoAを外してカルニチンをつける
アシルCoAは、ミトコンドリアに入る時に、一度CoAを外し、カルニチンと結合します。ミトコンドリアに入ると、カルニチンを外し、またCoAと結合し、アシルCoAに戻ります。

ミトコンドリア
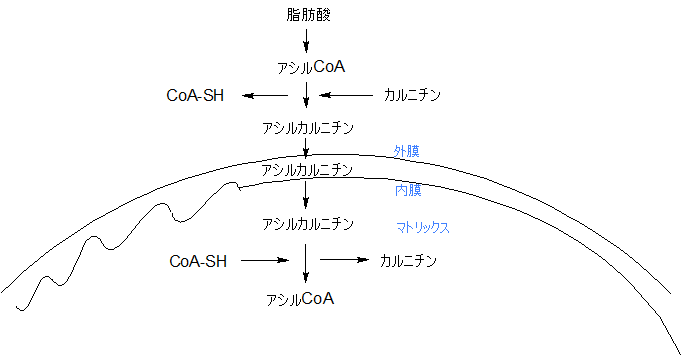
ミトコンドリアは図のような構造になっています。アシルCoAは、これからミトコンドリアのマトリックス内に移動します。しかし、アシルCoAはストレートにマトリックス内に入れないのです。
アシルCoAミトコンドリアへ
ミトコンドリアに入る前に、アシルCoAは、CoAを外し、カルニチンと結合してアシルカルニチンになり、ミトコンドリアの外膜と内膜を通過して、マトリックス内に移動します。
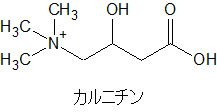
カルニチンはこんな構造式です。補酵素A(CoA)と比べるとだいぶ小さいです。
アシルカルニチンの一例として、炭素数16のパルミチンCoAを取りあげます。
パルミチンCoAがCoAを離してL-カルニチンと結合し、L-パルミトイルカルニチンになる反応を書きました。(出典)

(画像をクリックすると大きくなります)
アシルカルニチンはカルニチンを外して再びCoAをつける
マトリックス内に来ると、アシルカルニチンは、カルニチンを離し、再びCoAと結合してアシルCoAとなります。
この反応は上に書いた反応式が逆向きになるだけです。
アシルCoAは、これから4つの反応を経てアセチルCoAをつくります。
アシルCoAからアセチルCoAを切り離すまで
アシルCoAからアセチルCoAを切り離すまで4つの行程があります。
ここから先は、KEGGの Fatty acid degradation(脂肪酸分解)の反応経路を読んで行きます。
アシルCoAのβ位に二重結合ができる
最初の反応でα炭素とβ炭素の間に二重結合が形成されます。α、βとは、もともとカルボキシ基だった炭素(C)の隣の炭素(C)から、α、β・・・と数えていきます。
α炭素とβ炭素は1個ずつ合計2個水素(H)を離し、FADはその水素をもらってFADH2になります。
FADH2は、細胞が使うエネルギーATP(アデノシン三リン酸)生成に関わります。別な反応で使われます。
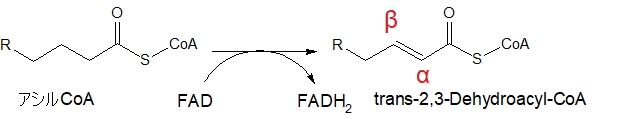
パルミチン酸にCoAが結合したパルミチルCoAの場合は、(E)-2-ヘキサデセノイルCoAになります。

(画像をクリックすると大きくなります)
以下、物質名はとりあえず無視していただいて、形の変化を見ていただくとよいと思います。
二重結合に水が反応してOHが付く
次に、酵素の反応でβ炭素の位置に形成された二重結合に水(H2O)が反応して、水酸化されます。
下の図で説明すると、α-β間の二重結合が解消され、β位に水酸基(-OH)が結合します。
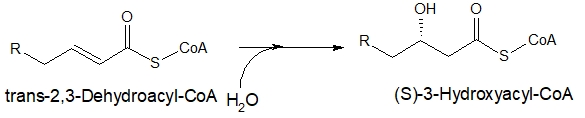
炭素数16のパルミチン酸から変化した(E)-2-ヘキサデセノイルCoAは、(S)-3-ヒドロキシヘキサデカノイルCoAになります。
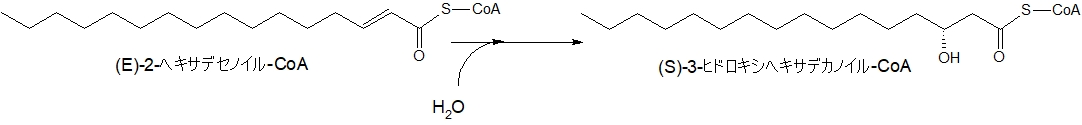
(画像をクリックすると大きくなります)
OHがケト基(=O)に変わる
さらに酸化されて、β炭素の位置にある水酸基(-OH)がケト基(=O)に変わります。
OHがついている炭素Cは、書いていませんが、もう1つ水素(H)と結合しています。その水素(H)と水酸基(-OH)についていた水素と合計2個が外れ、NAD+に供給されます。
NAD+は、NADH+H+になります。酸素は炭素と二重結合するように変わります。
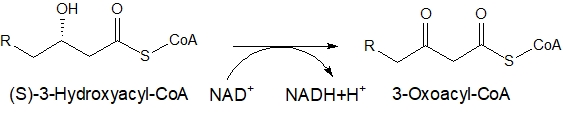
炭素数16の(S)-3-ヒドロキシヘキサデカノイルCoAは、3-オキソパルミトイル-CoAになります。
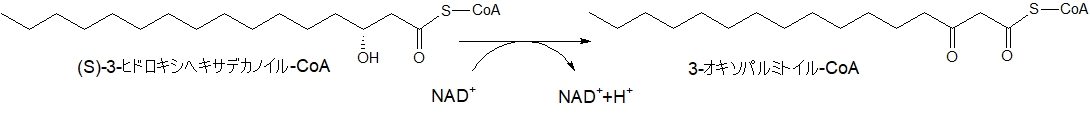
(画像をクリックすると大きくなります)
新たにHS-CoAが来てアセチルCoAが切り離される
さて、この最後の行程で、もともとのアシルCoAから炭素数2のアセチルCoAが切り離されます。
図を見ていただくとわかりやすいです。ちょうど2個ある酸素(O)の間で切れて、アセチルCoAを切り離し、残ったアシル基にCoAが結合して、炭素数が2個減った(短くなった)アシルCoAになります。
さて、炭素鎖が炭素2個分短くなった脂肪酸アシルCoAは、次のβ酸化の第1段階の基質となり、脂肪酸アシル鎖の部分がすべてアセチルCoA に酸化されるまで反応は繰り返されていきます。
炭素2個ずつ、一枚ずつ皮をはがされるように、脂肪が分解されていく感じでしょうか。
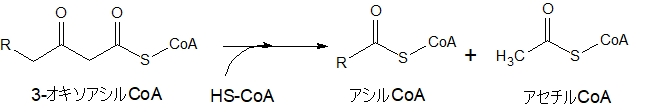
そして、このアセチルCoAは、TCA回路に入り、ATPをつくる反応に関与していきます。糖も脂肪もエネルギーをつくるときは、アセチルCoAとなり、TCA回路に入ります。
もともとパルミチン酸だった3-オキソパルミトレイルCoAの場合は、アセチルCoAを切り離し、炭素数14のミリストイルCoAとなります。

(画像をクリックすると大きくなります)
NOTE
β酸化は、脂肪酸から炭素2個ずつアセチルCoAを切り出す反応でしたが、逆に、アセチルCoAから脂肪酸をつくることができます。
ブドウ糖を脂肪酸に変えるという記事で書いています。
このときの反応とβ酸化の反応を見比べていただきたいのですが、反応の方向は正反対になりますが、変化の行程は同じです。
反応式だけを追っているとくたびれるかもしれませんが、最初に二重結合ができて、そこにOHがついて、さらにOになり、補酵素A(CoA)がついて、アセチルCoAを切り離すと知っておくとよいと思います。
脂肪酸が4つの行程で、アセチルCoAをつくり出すのは、ブドウ糖からアセチルCoAになるまでの反応経路が11行程あるのと比べると、とても簡単です。
脂肪酸とグリセリンからできた脂肪が、エネルギーをためこみ、また使うために一番適しているのは間違いありません。
脂肪をエネルギーにする他の記事は、脂肪をエネルギーにするをご覧下さい。